1902 Encyclopedia > Animal Kingdom > Animal Kingdom - The Metazoa
Animal Kingdom
(Part 3)
II. THE METAZOA
The germ becomes differentiated into histogenetic cells, and these cells become arranged into two sets, the one constituting the outer wall of the body, or extoderm, while the other, or endoderm, lies internal to the foregoing, and constitutes the lining of the alimentary cavity, when, as in usually the case, a distinct alimentary cavity exists. In the embryo, the representatives of these two layers are the epiblast and hypoblast.
All the Metazoa, in fact, commence their existence in the form of an ovum, which is essentially a nucleated cell, supplemented by more or less nutritive material, or food yelk. The ovum, after impregnation, divides into cleavage masses, or blastomeres, giving rise to a Morula, in the midst of which arises a cavity, the blastocoele (cleavage cavity, "Furchungshohle" of the Germans), which may be larger or smaller, filled only with fluid, or occupied by food yelk. When it is largest, the blastomeres, disposed in a single later, form a spheroidal vesicle, enclosing a correspondingly shaped blastocoele. When it is reduced to a minimum, the Morula is an almost solid aggregation of blastomeres, which may be nearly equal in size, or some may be much larger than others, in consequence of having undergone less rapid division. The next stage in the development of the embryo of a Metazoon consists (in all cases except a few parasitic anenterous forms) in the conversion of the Morula into a body having a digestive cavity, or a Gastrula. The animals in which the embryo takes on the form of a Gastrula, may be termed, as Haeckel has proposed, Gastroece.
The conversion of the Morula into the Gastrula may take place in several ways.
In the simplest, the Morula being composed of equal or nearly equal blastomeres, more or less completely converted into cells, these differentiate themselves into an outer layer, the epiblast, investing the remaining cells, which constitute the hypoblast. The central cells of the hypoblast next diverge and give rise to space filled with fluid, the alimentary cavity, which opens at one end, and thus gives rise to the Gastrula. This is the process generally observed in Porifera, Coelenterata, Turbellaria, Trematoda, and Nematoidea.
In a second class of cases, the Morula becomes converted into blastomeres of unequal sizes, a small and a large set. The smaller rapidly become converted into cells, and invest the larger and any remains of the food yelk, as a blastoderm, which at first represents only the epiblast of the former case. The hypoblast arises either from the epiblast thus formed, or from the included larger blastomeres. This is the process observed in certain Turbellaria, in the Ctenophora, in the Oligochoeta and Hirudinae, in the Arthropoda, and in most Vertebrata.
In a third group of instances, the Morula, whether consisting of equal or unequal blastomeres, becomes spheroidal, and encloses a correspondingly shaped blastocoele. One part of the wall of this vesicular Morula then becomes invaginated, and gives rise to the alimentary cavity, with the hypoblast which limits it. This process has been observed in the Choetognatha, Echinodermata, Gephyrea, polychaetous Annelida, Enteropneusta, Brachiopoda; in most Mollusca; in Amphioxus; and, slightly modified, in Petromyzon and in the Amphibia. These various modes in which the two primary layers of the germ may be developed shade off into one another, and do not affect the essence of the process, which is the segregation of one set of cells to form the external covering of the body, and of another set to constitute the lining of the alimentary canal.
In whatever manner the Gastrula is formed, and whatever be its shape when its alimentary cavity is complete, one of two things happens to it. It becomes provided with many ingestive apertures, distinct from that first formed; or with only one, which may or may not be distinct from the first aperture. The former division comprises only the Sponges (Porifera or Spongida) in which, as the remarkable researches of Haeckel have shown, the walls of t he deeply cu-shaped Gatrula become perforated by the numerous inhalent ostioles, while the primitive opening serves as the exhalent aperture. These may be termed the Metozoa polystomata.
The latter division includes all the remaining forms, which may be grouped together as Metazoa monostomata. Among these, two primary groups are distinguishable, of which the second exhibits an advance in organization upon the first. In the first, the aperture of the Gastrula becomes the permanent mouth (Archoeostomata). In the second, the permanent mouth is a secondary perforation of the body wall (Deuterostomata).
1. It is now well established that the aperture of the Gastrula becomes the oral aperture of the adult in the Coelenterata, which group includes animals differing much in grade of organization, from the simple Hydra to the complex Cteniphore, but all manifestly exhibiting variations of one fundamental type. Parallel with these may be ranged an assemblage composed of the Turbellaria, Rotifera, and Trematoda, which are associated together by the closet resemblances of structure, and which present an even greater range in grade of organization than the Coelenterata. The lower Rhadocoela come very close to the Infusoria (as close as the multicellular to the unicellular (Algae), and are but little superior to Hydra in the degree of their organic diffeentiation; while in the Trematoda, the land Planarioe, and the Nemertidoe, we have animals which Attain a considerable complexity, and in the case of many Trematoda and of Lineus (Pilidium) undergo remarkable metamorphoses. As a cognate group, the Nematoidea may be enumerated, extremely simple in their lowest forms, considerably differentiated in the higher, and connected with the Turbellaria by such forms as Polygordius. The Oligochoeta and the Hirudinea also belong to this division of Scolecimorpha, but they differ from the foregoing in the development of a segmented mesoblast.
In the Coelenterata, Nematoidea, Turbellaria, Trematoda, and Rotifera, the mode of origin of the cells which lie between the epiblast and the hypoblast, constitute the mesoblast, and give rise to the connective tissue and muscles of the body was wall and of that of the intestine, is not precisely known. They may take their origin in the epiblast, or in the hypoblast, or in both. But in the Oligochoeta and the Hirudinea, after the epiblast and hypoblast are differentiated, the cells of the latter give rise by division to two hands of cells, which lie, one on each side of the long axis of the ventral face of the worm, and constitute the mesoblast. This becomes marked out by transverse constriction into segments, and, in each segment, gives rise to all the tissues which lie between the epiblast (epidermis) and hypoblast (epithelium of the alimentary canal). The mouth corresponds with the primitive involution of the Mrotula; the anal aperture is a new formation. In the Nematoidea and in the lower Rhabdocoele Turbellaria, the intestinal calan is a simple tube or sac. But in some Turbellaria and trematoda, the alimentary canal gives off diverticula, which ramily through the mesoblast and even unite together. The like takes place in a great many Coelenterata, and the "gastrovascular apparatus," as it has been well termed, which is thus formed, is highly characteristic of them. The animals just referred to, therefore, have an "enterocoele" more or less distinct from the proper digestive cavity, but connected with it, and ramifying through the mesoblast.
2. In the remaining members of the animal kingdom, the embryo develops a secondary mouth as a perforation of the body wall, the primary aperture sometimes becoming the anus, and sometimes disappearing. Of these Metazoa deuterostomata, there are some which follow the mode of development of the Oligochoeta and Hirudinea very closely, so far as the formation and segmentation of the mesoblast is concerned, though the question whether this segmented mesoblast arises from the epiblast or the hypoblast, has not been exhaustively worked out. These are the Annelida polychoeta; and thre is the closest resemblance in development between them and the lower Arthropoda (Crustacea, Arachnida, lower Insecta), while, in the higher Arthropods, the process is complicated by the development of an amnion, and by some other special peculiarities which need not be considered in detail. In all these Metazoa, whatever cavities are developed in the mesoblast, whether a wide perivisceral cavity, or vascular canals, or both combined, they arise from the splitting or excavation of the mesoblast itself, and are not prolongations of the alimentary cavity. Hence they may be termed Schizocoela.
But, in certain other deuterostomatous Metazoa, the mesoblast becomes excavated, and a "perivisceral cavity" and vessels are formed in quite another gashion.
Thus in the Choetognatha, represented by the strange and apparently anomalous Sagitta, Kowalewsky's researches show, that the vitellus undergoes complete segmentation, and is converted into a vesicular Morula, on one side of which involution takes place, and give rise to the primitive alimentary canal, of which the opening of involution becomes the permanent anus, the mouth being formed by perforation at the opposite end of the body. Before the mouth is formed, however, the primitive alimentary cavity throws out, on each side, a caecall pouch, which extends as far forwards as its central continuation does, and grows backward behind the anus. The two sacs thus prolonged posteriorly, meet, but remain divided from one another by their applied walls in the median line. These lateral sacs now become shut off from the median portion of the primitive alimentary cavity (which opens at its anterior end, and becomes the permanent alimentary canal), and are converted into shut sacs, the cavity of each of which forms one-half of the perivisceral cavity. The inner wall of each sac, applied to the hypoblast, gives rise to the muscular wall of the intestine; and the outer wall, applied to the epiblast, becomes the muscular wall of the body, and from it the generative organs are evolved. The great ganglia and nerves are developed from the cells of the epiblast. Thus Sagitta is temporarily coelenterate, but the two gastrovascular sacs, each enclosing an entercoele, become shut off from the alimentary canal and metamorphosed into the walls of the perivisceral cavity. But it is not altogether clear whether the cells of the enterocoele give rise only to the lining of the perivisceral cavity, and whether the muscles and connective tissue are otherwise derived or not. Kowalesky's evidence, however, is in favour of the origin of the muscles directly from the cells of the mesoblastic diverticula.
In the Echinodermata, the brilliant investigation of Johannes Muller, confirmed in their general features by all subsequent observers, proved, firstly, that the ciliated embryonic Gastrula (the primitive alimentary canal of which is formed by involution of a vesicular blastoderm), to which the egg of all ordinary Echinoderms give rise, acquires a mouth, by the formation of an aperture in the body wall, distinct from the primitive aperture of the gastrula, so that, in this respect, it differs from the embryo of all Coelenterata; secondly; that the embryo thus provided with mouth, stomach, intestine, and anus acquires a complete bilateral symmetry; thirdly, that the cilia, with which it is primitively covered, are ultimately restricted to one or more series, some of which encircle the axis of the body, or a line drawn from the oral to the anal apertures; and fourthly, that, within this bilaterally symmetrical larva or echinopoedium, as it may be called, the more or less completely radiate Echinoderm is developed by a process of internal modification.
Muller believed that the first step in this process was the ingrowth of a diverticulum of the integument, as a hollow process, which became converted into the ambulacral vascular system of the Echinoderm. He did not attempt to explain the origin of the so-called blood-vascular sys tem, or pseudhaemal vessels, nor the perivisceral cavity, Muller's conclusions remained unchallenged until 1864. when Prof. Alexander Agassiz took up the question afresh, and, in a remarkable paper on the development of the genus Asteracanthion, detailed the observations which led him to believe that the ambulacral vessels do not arise by involution of the external integument, but that they commence as two primitively symmetrical diverticula of the stomach (the "wursformige Korper" of Muller), one of which becomes connected with the exterior by an opening (the forsal pore observed by Muller, and considered by him to be the origin of the ambulacral vessels), and gives rise to the ambulacral vessels, the ambulacral region of the body of the Ecginoderm being modeled upon it; while upon the othr gastric sac, the antambulacral wall of the starfish body is similarly modeled. Both gastric sacs early become completely separated from the stomach of the echinopoedium, and open into one another, so as to from a single horseshoe-shaped sac, connected with the exterior by a tube which is converted into the madreporic canal. Agassiz does not explain the mode of formation of the perivisceral cavity of the starfish, and has nothing to say respecting the origin of the pseudhaemal vessels.
Recently, Metschnikoff has confirmed the observations of Aggassiz, so far as the development of the ambulacral system from one of the diverticula of the alimentary canal of the starfish larva is concerned, and he has added the important discovery that the perisvisceral cavity of the echinoderm is the product of what remains of these diverticula. Moreover, his observations on other Echinodermata show that essentially the same process development of the peritoneal cavity occurs in Ophiuridoe, Echinidoe, and Holothuridoe.
The precise mode of origin of the pseudhaemal system, or so-called blood-vessels, of the echinoderms, is not yet made out. But it is known that the cavity of these vessels contains corpuscles similar to those which are found in the perivisceral cavity and in the ambulacral vessels, and that all three communicate.
Both Agassiz and Metschnikoff justly insist upon the correspondence in development of the lateral gastric diverticula of the Echinopoedium with the gastrovascular canal system of the Ctenophora; and on the ground of this resemblance, the former refers the echinoderms to the Radiata, retaining under that Cuvierian denomination the Acalephoe (Coelenterata) and the Echinodermata. But this arrangement ignores the real value of his own discovery, which shows that the Echinoderms have made a great and remarkable step, in passing from their primarily coelenterate stage of organization to their adult condition. And it further ignores the unquestionable fact, admirably brought out by the same excellent observe's recent investigations into the development of Balanoglossus, that the Echinopoedium is almost identical in structure with young of animals, such as the Gephyrea and Enteropneusta, which are in no sense radiate, but are, eminently, bilaterally symmetrical. In fact, the larva of Balanoglossus (the sole representative of the Enteropneusta) was originally described by Muller, under the name of Tornaria, as an Echinoderm larva; and was subsequently more fully examined by Prof. Alex Agassiz, who also regarded it as an unquestionable Echinoderm larva; and it is only recently that it has been proved, partly by Metschnikoff and partly by Aggasiz himself, to be the larval form of Balanoglossus. In Balanoglossus, as in the echinoderms, it appears that saccular outgrowths of the intestine give rise to the perivisveral cavity and its walls; and, if such be the case, thes mesoblast will be chiefly, if not wholly, represented by diverticula of the alimentary canal. Thus in the Choetognatha and echinodermata, and possibly in the Enteropneusta, the perivisceral cavity is a portion of the alimentary cavity shut off from the rest; and, in contradistinction to the Schizocoela, in which the perivisceral cavity is produced by a splitting of the mesoblast, they may be said to be Enterocoela.
If we endeavour to determine the place of the three remaining great groups of animals, the Mollusca, the Tunicata, and the Vertebrata, obstacles arise, - firstly, from a want of sufficiently exact knowledge respecting the Mollusca; and secondly, from the difficulty of interpreting certain well-ascertained facts in the Vertebrata.
That the Mollusca, including under that name the Polyzoa and Brachiopoda, as well as the higher Mollusks, are closely allied to the annelida, is readily demonstrated. The known forms of Brachiopod, Lamellibranch, Pteropod, and Gasteropod larvae all have their parallels among Annelidan larvae. The Polyzoa are closely allied with the gephyrea and Rotifera; and a Mollusk may be said to be a few-segmented annelid with a mantle. But whether the perivisceral cavity is developed in the annelidan or in the echinoderm fashion is not yet clear. In the Polyzoa, the evidence is at present insufficient to justify any conclusion. In the Brachiopoda, there is some ground for thinking that the perivisceral cavity is formed in the same way as in Sagitta and the Echinodermata; while, in the Lamellibranchiata and Odontophora, there is every reason to believe that the perivisceral cavity is formed by splitting of the mesoblast, or that they are schizocoelous.
In the lowest Tunicata, represented by Appendicularia, the recent investigations of Fol have shown, that, in the adult, the body proper is formed almost exclusively by an ectoderm and endoderm, which proceed directly from the epiblast and hypoblast of the embryo. It is only in the caudal appendage that a distinct mesoblast is represented by the notochord and the muscles. The blood channels correspond with the blastocoele, and the "house" in which these singular animals shelter themselves is a cuticular secretion, representing the cellulose coat of the higher ascidians. The Appendicularioe have no atrium, or at most only rudiments of it, hence the branchial clefts open directly on the haemal aspect of the body, which corresponds with the ventral face of a vertebrate animal. In all other Tunicata, and atrial cavity is formed by involution of the ectoderm, which thus give rise to a cavity on each side of the branchial sac, into which the branchial clefts of the adult open; and a thick cellulose cuticula, into which cells from the ectoderm usually wander, invests the exterior of the body. The "atrial tunic," or invaginated layer of the ectoderm, is reflected, as a visceral layer, over more or less of the outer surface of the alimentary canal, and, as a parietal layer, over more or less of the inner surface of the body wall; and the space between the two (the blastocoele) becomes converted into the blood passages. Thus, such an ascidian resembles a vertebrated animal, not only in the manner in which its nervous centre is developed, but in the fact that it possesses an atrial cavity, which singularly resembles the pleuroperitoneal chamber of a vertebrate. For this cavity is bounded externally by the atrial tunic and the integument, which correspond with the somatopleural layer of the mesoblast and the epiblast of a vertebrate embryo; and it is bounded, internally, by the atrial tunic and the epithelium of the alimentary canal, which, to the same extent, correspond with the splanchnopleure and the hypoblast. The primitively double atrial aperture has its parallel in the peritoneal openings which persist in many Vertebrata.
Thus the ascidian has no "perivisceral cavity" formed by splitting of the mesoblast, nor has it any "perivisceral cavity" formed by diverticula from the alimentary canal. It is neither enterocoelous nor schizocoelous, but what, at first sight, resembles a perivisceral cavity is formed within the body by involution, and the ascidian may therefore be said to be epicoelous. If the alate prolongations of the body which lie at the sides of the branchial apertures, in Balanoglossus, were to enlarge and unite round the anus so as to leave but a relatively small opening between their edges, the cavity so formed would answer to the atrial chamber of an ascidian.
In the higher Vertebrata, the pleuroperitoneal1 cavity appears to be formed by the splitting of the mesoblast into two layers, a splanchnopleure and a somatopleure, and, therefore, seems at first to correspond with the perivisceral cavity of the Annelids and Arthopods. But what is now known of the structure and development of the lowest and most embryonic of known Vertebrata, Amphioxus, throws very great doubt upon this interpretation of the facts. One of the most singular of the many peculiarities of Amphioxus is the fact that the branchial clefts open, not on the exterior of the body, as in all other Vertebrata, but into a chamber with a single external aperture, which, on the one hand, curiously resembles the atrium of an ascidian; while, on the other, it is undoubtedly homologous with the pleuroperitoneal cavity of the higher Vertebrata. Now Kowalesky's investigations have shown, that, at first, the branchial apertures of the embryo Amphioxus open upon the exterior of the body, but that, after a time, a process of the wall of the body, on the dorsal side of the branchial apertures, grows down over them, and, uniting with its fellow in the median ventral line of the body at all points, except at the abdominal pore, gives rise to the outer wall of the pleuroperitoneal cavity. Thus the lining of that cavity, like the atrial tunic of the ascidian, is a derivative of the epiblast; and Amphioxus is epicoelous. As it can hardly be doubted that the somatopleure of Amphioxus is the homologue of the somatopleure in the higher Vertebrata, it becomes highly probable that the apparent splitting of the mesoblast in the latter, after all, represents the mode of development of the pleuroperitoneal cavity which obtains in the former, and, thus, that the Vertebrata are not schizocoelous, but epicoelous. Whether this suggestion will turn out to be well based or not, must be decided by the embryological investigations specially directed to this point: but that there should be any essential difference between Amphioxus and other Vertebrata, in the manner in which the pleuroperitoneal cavity is formed, is highly improbable.
The distance between Amphioxus and other vertebrate animals, which has hitherto been generally supposed to exist, has been greatly diminished by recent investigations. So far from being devoid of a brain and of a skull, the regions of the cerebro-spinal axis and of the neural canal, which answer to those organs in the higher Vertebrata, are, in proportion, extremely long in Amphioxus, as they are in all vertebrate embryos. But, in Amphioxus, the head retains throughout life a segmentation comparable to that of the rest of the body, while, in the higher Vertebrata, almost all traces of these distinct segments are very early lost. Moreover, in Amphioxus, the renal apparatus, so far from being absent, is represented by a comparatively large structure, and nothing is wanted to equip it with all the organs found in a young Marsipobranch, but auditory sacs, which, however, it must be remembered, make their appearance late in the Lamprey. With all this, the gap between Amphioxus and the Marsipobranchii is undoubtedly more considerable than that between the Marsipobranchii and other fishes, and it may represent a primary division of the class Pisces, - which, from the segmentation of the skull, may be termed the Entomocrania, - as opposed to the rest, in which the primary segmentation of the skull is almost completely effaced, and which may therefore be designated Holocrania.
It has been stated above that the great majority of the Metazoa pass through the Gastrula condition, and belong to the division of the Gastroeoe. In some members of this division, however, the alimentary canal may be rudimentary, as in sundry male Rotifera and in the Gordiacei among the Nematoidea, and yet these are so closely allied to other forms possessing fully developed digestive canals, that it is reasonable to regard their rudimentary alimentary apparatus as absorbed. In two groups, however, the Cestoidea and the Acanthocephala, there is no trace of an alimentary canal either in the embryo or in the adult.
From the point of view of phylogeny, this fact may be interpreted in two ways. Either the alimentary canal which once existed has aborted, and the Cestoidea and Acanthocephala are modified Scolecimorpha, or these parasites have not descended from Gastroeoe, but have passed into their present condition directly form a Morula-like form of Metazoon. In the latter case they will from a division of Agastroeoe, apart from the other Metazoa.
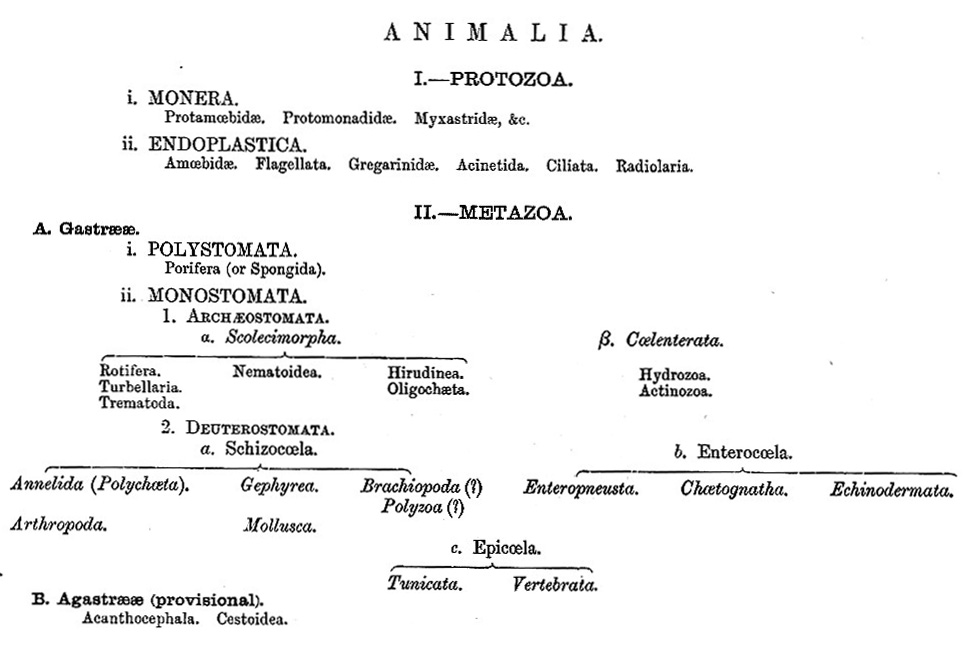
The subjoined synopsis indicates the general relations of the different groups of the Animal kingdom, in accordance with the views which have been put forward in the preceding pages.
Those who are familiar with the existing condition of our knowledge of animal morphology, will be aware that any such scheme must needs, at present, be tentative and subject to extensive revision, in correspondence with the advance of knowledge. Nor will they regard it as any objection to the scheme of classification proposed, that the divisions sketched out may be incapable of sharp definition-the constant tendency of modern investigations being to break through all boundaries of groups, and to fill up the gaps between them by the discovery of transitional forms. In the place of assemblages of distinctly definable groups, which it has hitherto been the object of the Taxonomist to define and co-ordinate in precise logical categories, we are gradually learning to substitute series, in which all the modifications by which a fundamental form passes from lower to higher degrees of organic complications, are summed up. (T. H. H.)
Read the rest of this article:
Animal Kingdom - Table of Contents
The above article was written by the Rt. Hon. Thomas Henry Huxley, M.D., LL.D., F.R.S., President of the Royal Society, 1883-85; Professor of Natural History in the Royal School of Mines, London, 1854; author of Theory of the Vertebrate Skull, The Physical Basis of Life, Introduction to the Classification of Animals, Lay Sermons, Elementary Physiology, etc.
|